Hva er loven om økologisk tiende eller 10%?
Loven om økologisk tiende, økologisk lov eller 10% øker måten energien beveger seg i ved avledning gjennom de forskjellige trofiske nivåene. Det er også ofte sagt at denne loven bare er en direkte følge av Termodynamikkens andre lov.
Den økologiske energien er en del av økologien som er opptatt av å kvantifisere forholdene vi har skissert ovenfor. Det er ansett at Raymond Lindemann (spesielt i hans seminalarbeid fra 1942) var den som etablerte grunnlaget for dette studiet.

Hans arbeid fokuserte på begreper kjede og trofiske nettverk, og på kvantifisering av effektivitet i overføring av energi mellom de forskjellige trofiske nivåene.
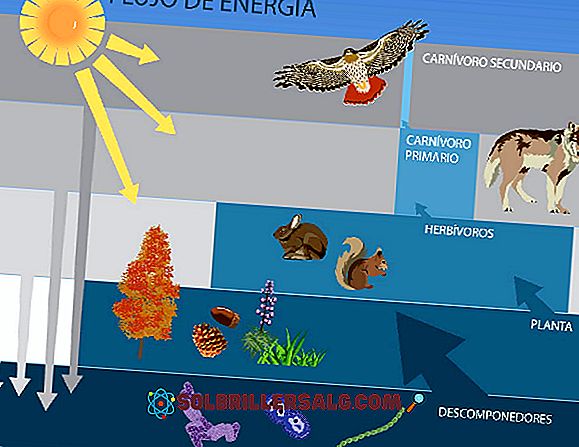
Lindemann starter fra hendelsen solstråling eller energi som et samfunn mottar, gjennom fangstene som gjøres av plantene gjennom fotosyntese, og fortsetter å overvåke fangsten og dens påfølgende bruk av herbivorene (primære forbrukere), deretter av kjøttetankene (sekundære forbrukere). ) og til slutt av dekomponere.
Hva er loven til den økologiske tienden?
Etter Lindemanns banebrytende arbeid ble det antatt at trofiske overføringseffektiviteter var rundt 10%; Faktisk refererte noen økologer til en 10% lov. Siden da har det blitt generert flere forvirringer angående dette problemet.
Det er absolutt ingen naturlov som resulterer nettopp i at en tiendedel av energien som går inn i et trofisk nivå, er det som overføres til det neste.
For eksempel viste en samling av trofiske studier (i marine og ferskvannsmiljøer) at overføringseffektiviteten ved trofisk nivå varierte mellom ca 2 og 24%, selv om gjennomsnittet var 10, 13%.
Som en generell regel, som er anvendelig for både akvatiske og jordbaserte systemer, kan det sies at sekundær produktivitet av herbivorer vanligvis ligger omtrent, en størrelsesorden under den primære produktiviteten som den er basert på.
Dette er ofte et konsistent forhold som opprettholdes i alle foraging-systemer, og som vanligvis skjer i strukturer av pyramidaltypen, hvor basen er tilveiebrakt av plantene, og på dette grunnlag er en mindre basert på de primære forbrukere, på hvilken en annen (mindre fremdeles) av sekundære forbrukere sitter.
Organisasjonsnivåer
Alle levende vesener krever materie og energi; Materiell for bygging av deres kropper og energi for å utføre sine vitale funksjoner. Dette kravet er ikke begrenset til en individuell organisme, men utvides til høyere nivåer av biologisk organisasjon som disse individene kan samsvare med.
Disse organisasjonsnivåene er:
- En biologisk populasjon : organismer av samme art som lever i samme bestemte område.
- Et biologisk samfunn : sett av organismer av forskjellige arter eller populasjoner, som bor i et bestemt område og samhandler gjennom mat eller trofiske forhold).
- Et økosystem : det mest komplekse nivået av biologisk organisasjon, utgjør et samfunn relatert til dets abiotiske miljøvann, sollys, klima og andre faktorer, som det samhandler med.
Trofiske nivåer
I et økosystem etablerer samfunnet og miljøet strømmer av energi og materie.
Organene i et økosystem er gruppert i henhold til en "rolle" eller "funksjon" som de oppfyller innen fordøyelses- eller trofiske kjeder; Slik snakker vi om trofiske nivåer av produsenter, forbrukere og dekomponere.
I sin tur virker hver og en av disse trofiske nivåene med det fysisk-kjemiske miljøet som gir forholdene for livet og samtidig fungerer som kilde og synke av energi og materie.
Grunnleggende begreper
Grov og netto primær produktivitet
Først må vi definere primærproduktivitet, som er hvor mye biomasse produseres per areal.
Det er vanligvis uttrykt i enheter av energi (Joules per kvadratmeter per dag), eller i enheter av tørr organisk materiale (kilo per hektar og per år), eller som karbon (masse karbon i kg per kvadratmeter per år).
Generelt, når vi refererer til all energi som er fastlagt ved fotosyntese, kalles vi vanligvis grov primærproduktivitet (PPG).
Fra dette blir en andel brukt på autotrofer selv (RA) respirasjon og er tapt i form av varme. Netto primærproduksjon (PPN) er oppnådd ved å subtrahere denne mengden fra PPG (PPN = PPG-RA).
Denne primære primærproduksjonen (PPN) er det som til slutt er tilgjengelig for forbruk av heterotrofer (det er bakterier, sopp og andre dyr som vi vet).
Sekundær produktivitet
Sekundær produktivitet (PS) er definert som produksjonshastigheten av ny biomasse av heterotrofe organismer. I motsetning til planter, heterotrofiske bakterier, sopp og dyr, kan de ikke gjøre de komplekse, energirike stoffene de trenger fra enkle molekyler.
De får sin materie og energi alltid fra planter, som de kan gjøre direkte ved å konsumere plantemateriale eller indirekte ved å mate på andre heterotrofer.
Det er på denne måten at fotosyntetiske planter eller organismer generelt (også kalt produsenter), omfatter det første trofiske nivået i et samfunn; De primære forbrukerne (de som lever på produsentene) utgjør det andre trofiske nivået og de sekundære forbrukerne (også kalt rovdyr) utgjør det tredje nivået.
Overføringseffektivitet og energiruter
Forholdet mellom netto primærproduksjon som strømmer langs hver av de potensielle energibaneene, er i siste instans avhengig av overføringseffektivitet, det vil si på hvilken måte energi blir brukt og beveger seg fra ett nivå til et annet. andre.
Effektkategorier for energioverføring
Det er tre kategorier med energieffektivitet, og med disse godt definerte kan vi forutsi mønstrene av energiflømning på trofiske nivåer. Disse kategoriene er: Effektiviteten av forbruk (EC), effektiviteten av assimilering (EA) og effektiviteten av produksjonen (EP).
La oss nå definere disse tre kategoriene nevnt.
Matematisk kan vi definere forbrukseffektiviteten (EC) på følgende måte:
EC = I n / P n-1 × 100
Der vi kan se at CE er en prosentandel av den totale tilgjengelige produktiviteten ( P n-1 ) som effektivt inntas av det øvre sammenhengende trofiske rommet ( I n ).
For eksempel, for primærforbrukere i beiteanlegget, er EC prosentandelen (uttrykt i enheter av energi og per tidsenhet) av PPN som forbrukes av plantelevende dyr.
Hvis vi refererte til sekundærforbrukerne, ville det være lik prosentandelen av produktiviteten til plantelevende dyr, forbrukes av kjøttetrærene. Resten dør uten å bli spist og går inn i dekomponeringskæden.
På den annen side uttrykkes effektiviteten av assimilering som følger:
EA = A n / I n × 100
Igjen refererer vi til en prosentandel, men denne gangen til den delen av energien som kommer fra maten, og inntas i et trofisk rom av en forbruker ( I n ) og som er assimilert av fordøyelsessystemet ( A n ).
Denne energien vil være tilgjengelig for vekst og for utførelse av arbeid. Rester (den ikke-assimilerte delen) går tapt med avføring og går deretter inn i trofisk nivå av dekomponeringene.
Til slutt uttrykkes produksjonseffektiviteten (PE) som:
EP = P n / A n × 100
som også er en prosentandel, men i dette tilfellet refererer vi til den assimilerte energien ( A n ) som ender opp med å bli innlemmet i ny biomasse ( P n ). Alle uassimilerte energiske rester går tapt i form av varme under respirasjon.
Produkter som sekresjoner og / eller utskillelser (rik på energi), som har deltatt i metabolske prosesser, kan betraktes som produksjon, Pn, og er tilgjengelige som lik, for nedbrytere.
Global overføringseffektivitet
Etter å ha definert disse tre viktige kategoriene, kan vi nå spørre om "global overføringseffektivitet" fra ett trofisk nivå til det neste, som bare er gitt av produktet av de nevnte effektivitetene ( EC x EA x EP ).
Uttrykt samtalende kan vi si at effektiviteten til et nivå er gitt av det som kan inntas effektivt, som deretter assimileres og ender opp med å bli innlemmet i ny biomasse.
Hvor går den tapte energien?
Produktiviteten av plantelevende dyr er alltid lavere enn den av plantene de matter fra. Vi kan da spørre: Hvor går den tapt energi?
For å svare på dette spørsmålet må vi trekke oppmerksomheten til følgende fakta:
- Ikke all biomasse av plantene forbrukes av plantelevende dyr, så mye av det dør og går inn i trofisk nivå av dekomponerer (bakterier, sopp og resten av detritivorer).
- Ikke all biomasse som forbrukes av herbivorene, eller den av herbivorene som konsumeres i sin tur av rovdyrene, blir assimilert og er tilgjengelig for å bli innlemmet i forbrukerens biomasse. en del går tapt med avføring og på den måten går den videre til dekomponeringene.
- Ikke all energien som kommer til å bli assimilert blir faktisk biomasse, siden en del går tapt i form av varme under åndedrettsvern.
Dette skjer av to grunnleggende grunner: For det første fordi det ikke er noen energikonverteringsprosess som er 100% effektiv. Det vil si at det alltid er et tap i form av varme i konvertering, som er i perfekt sammenheng med den andre loven om termodynamikk.
For det andre, siden dyr trenger å utføre arbeid, som krever energiforbruk og i sin tur innebærer nye tap i form av varme.
Disse mønstrene forekommer på alle trofiske nivåer, og som forutsatt av den andre loven om termodynamikk, deles en del av energien som forsøks å overføre fra ett nivå til et annet, alltid i form av varme som ikke kan brukes.