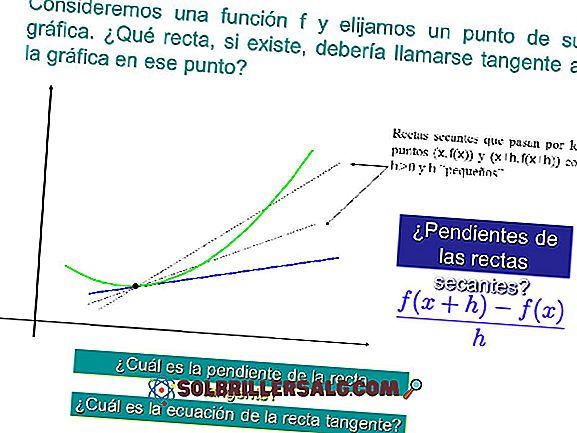
Transposons: typer og egenskaper
Transposons eller transponable elementer er fragmenter av DNA som kan endre deres plassering i genomet. Hendelsen for bevegelse kalles transposisjon og kan gjøre det fra en stilling til en annen, innenfor samme kromosom, eller endre kromosomet. De er til stede i alle genomene, og i betydelig grad. De har blitt grundig studert i bakterier, i gjær, i Drosophila og i mais.
Disse elementene er delt inn i to grupper, med tanke på elementets transposisjonsmekanisme. Dermed har vi retrotransposons som bruker et RNA-mellomprodukt (ribonukleinsyre), mens den andre gruppen bruker et DNA-mellomprodukt. Denne siste gruppen er transposones sensus stricto.
En nyere og detaljert klassifisering bruker den generelle strukturen av elementene, eksistensen av lignende motiver og identiteten og likhetene til DNA og aminosyrer. På denne måten defineres underklasser, superfamilier, familier og underfamilier av transponeringselementer.
Historisk perspektiv
Takket være undersøkelsen utført på mais ( Zea mays ) av Barbara McClintock i midten av 1940-tallet, kunne den tradisjonelle oppfatningen at hvert gen hadde et fast sted på et bestemt kromosom og fikset i genomet, bli endret.
Disse forsøkene gjorde det klart at enkelte elementer hadde evnen til å endre posisjon, fra ett kromosom til et annet.
Opprinnelig utpekte McClintock begrepet "kontrollerende elementer", siden de kontrollerte uttrykket av genet der de ble satt inn. Derefter ble elementene kalt hoppende gener, mobile gener, mobile genetiske elementer og transposoner.
I lang tid ble dette fenomenet ikke akseptert av alle biologer, og ble behandlet med litt skepsis. I dag er mobilelementer fullt akseptert.
Historisk sett ble transposoner betraktet som segmenter av "egoistisk" DNA. Etter 80-tallet begynte dette perspektivet å endres, siden det var mulig å identifisere interaksjonene og virkningen av transposonene i genomet, fra strukturell og funksjonell synspunkt.
Av disse grunner, selv om elementets mobilitet kan være skadelig i visse tilfeller, kan det være fordelaktig for populasjoner av organismer - analog med en "nyttig parasitt".
Generelle egenskaper
Transposoner er diskrete fragmenter av DNA som har evnen til å bevege seg innenfor et genom (kalt "vertsgenomet"), som vanligvis lager kopier av seg selv under mobiliseringsprosessen. Forståelsen av transposons, deres egenskaper og deres rolle i genomet, har endret seg gjennom årene.
Noen forfattere anser at et "transponabelt element" er et paraplybetegnelse for å betegne en serie gener med forskjellige egenskaper. De fleste av disse har bare den nødvendige sekvensen for deres gjennomføring.
Selv om alle deler karakteristikken for å kunne bevege seg gjennom genomet, kan noen legge igjen en kopi av seg selv på det opprinnelige stedet, noe som fører til økningen av transponeringselementer i genomet.
overflod
Sekvenseringen av forskjellige organismer (mikroorganismer, planter, dyr, blant andre) har vist at transponeringselementer eksisterer nesten i alle levende vesener.
Transposons er rikelig. I genomene av vertebrater okkuperer de fra 4 til 60% av alt genetisk materiale av organismen, og i amfibier og i en viss gruppe fisk, er transposonene ekstremt varierte. Det er ekstreme tilfeller, som mais, hvor transposoner utgjør mer enn 80% av genomet av disse plantene.
Hos mennesker er transponeringselementer ansett som de mest omfattende komponentene i genomet, med en overflod på nesten 50%. Til tross for deres bemerkelsesverdige overflod, har ikke rollen de spiller på genetisk nivå blitt fullstendig uttalt.
For å lage denne komparative figuren, la oss ta hensyn til de kodende DNA-sekvensene. Disse transkriberes til et messenger-RNA som endelig blir oversatt til et protein. I primater dekker kodende DNA bare 2% av genomet.
Typer transpononer
Vanligvis klassifiseres transponable elementer i henhold til måten de mobiliseres av genomet. På denne måten har vi to kategorier: elementene i klasse 1 og klasse 2.
Elementer i klasse 1
De kalles også RNA-elementer, fordi DNA-elementet i genomet transkriberes i en kopi av RNA. Deretter konverteres RNA-kopien tilbake til et annet DNA som settes inn i målstedet til vertsgenomet.
De er også kjent som retro-elementer, siden deres bevegelse er gitt av den omvendte strømmen av genetisk informasjon, fra RNA til DNA.
Antallet av denne typen elementer i genomet er enormt. For eksempel, Alu- sekvensene i det humane genom.
Transposisjonen er av replikativ type, det vil si at sekvensen forblir intakt etter fenomenet.
Elementer av klasse 2
Elementene i klasse 2 er kjent som DNA-elementer. I denne kategorien er transposoner som beveger seg fra ett sted til et annet uten behov for en mellommann.
Transposisjonen kan være av replikativ type, som i tilfelle av elementene i klasse I, eller det kan være konservativ: elementet er delt i hendelsen, slik at antall transponeringselementer ikke øker. Varene som ble oppdaget av Barbara McClintock tilhørte klasse 2.
Hvordan påvirker transposisjon verten?
Som vi nevnte, er transposoner elementer som kan bevege seg innenfor samme kromosom, eller hoppe til en annen. Imidlertid må vi spørre hvordan individets trening påvirkes på grunn av transponeringshendelsen. Dette avhenger hovedsakelig av regionen der elementet er transponert.
Dermed kan mobilisering påvirke verten positivt eller negativt enten ved å inaktivere et gen, modulere genuttrykk eller indusere uønsket rekombination.
Hvis vertenes kondisjon er drastisk redusert, vil dette faktum få effekt på transposonet, siden organismenes overlevelse er kritisk for dens vedvarende virkning.
Av denne grunn har visse strategier blitt identifisert i verten og i transposon som bidrar til å redusere den negative effekten av transposisjonen, å oppnå balanse.
For eksempel må noen transposoner settes inn i regioner som ikke er essensielle i genomet. Dermed påvirker virkningen sannsynligvis minimal serie, som i heterochromatin-områdene.
På vertsdelens side inkluderer strategier DNA-metylering, som reduserer ekspresjonen av det transponerbare element. I tillegg kan noen forstyrrende RNAs bidra til denne oppgaven.
Genetiske effekter
Transposisjon fører til to grunnleggende genetiske effekter. For det første forårsaker de mutasjoner. For eksempel er 10% av alle genetiske mutasjoner i musen et resultat av transponeringer av retro-elementer, hvorav mange er kodende eller regulatoriske regioner.
For det andre fremmer transposoner hendelser av uønskede rekombinasjoner, som resulterer i omkonfigurering av gener eller hele kromosomer, som generelt bærer med dem deletjoner av det genetiske materialet. Det er anslått at 0, 3% av genetiske lidelser hos mennesker (som arvelige leukemier) oppsto på denne måten.
Det antas at reduksjonen av vertskapstrening på grunn av skadelige mutasjoner er hovedgrunnen til at transponeringselementene ikke er mer rikelig enn de allerede er.
Funksjoner av transponeringselementene
Opprinnelig ble det antatt at transposoner var parasittergener som ikke hadde noen funksjon i sine verter. I dag, takket være tilgjengeligheten av genomiske data, har det blitt lagt større vekt på sine mulige funksjoner og til transposons rolle i utviklingen av genomene.
Noen formodede regulatoriske sekvenser er avledet fra transponable elementer og har blitt bevart i flere hvirveldyrsleder, i tillegg til å være ansvarlig for flere evolusjonære nyheter.
Rolle i utviklingen av genomene
I følge nylig forskning har transposoner hatt en betydelig innvirkning på arkitekturen og utviklingen av genomene av organiske vesener.
I en liten skala er transposons i stand til å formidle endringer i bindingsgrupper, selv om de også kan ha mer relevante virkninger som betydelige strukturelle endringer i genomisk variasjon, slik som deletjoner, duplikasjoner, inversjoner, duplikasjoner og translokasjoner.
Det antas at transposoner har vært svært viktige faktorer som har formet størrelsen på genomene og deres sammensetning i eukaryote organismer. Faktisk er det en lineær korrelasjon mellom størrelsen på genomet og innholdet av transponeringselementer.
eksempler
Transposons kan også føre til adaptiv utvikling. De klareste eksemplene på transposons bidrag er utviklingen av immunsystemet og transkripsjonsreguleringen via ikke-kodende elementer i moderkaken og i hjernen til pattedyr.
I immunsystemet til vertebrater produseres hvert av det store antall antistoffer ved hjelp av et gen med tre sekvenser (V, D og J). Disse sekvensene er fysisk separert i genomet, men de kommer sammen under immunresponsen gjennom en mekanisme kjent som VDJ rekombination.
På slutten av 1990-tallet fant en gruppe forskere at proteinene som var ansvarlige for VDJ-binding, ble kodet med RAG1- og RAG2- gener . Disse manglet introner og kunne forårsake transponering av bestemte sekvenser i DNA-mål.
Mangelen på introner er et vanlig trekk ved genene som er avledet av retrotransposisjon av et messenger-RNA. Forfatterne av denne studien antydet at immunsystemet til vertebrater oppsto takket være transposoner som inneholdt forfedre til RAG1- og RAG2- gener .
Det er anslått at ca. 200 000 innsettinger har blitt uttrykt i slaktingen av pattedyr.